






 Հիստոհամատեղելիությունը MHC-ն գեների մի խումբ է,որի արգասիքները կարևոր դեր են խաղում իմուն պատասխանի կայացման և սեփականը օտարից տարբերելու գործում: Կոչվում են հիստոհամատեղելիության գեներ,քանի որ նրանցով է որոշվում,թե փոխպատվաստված հյուսվածքը կընդունվի տիրոջ կողմից (հիստոհամատեղելի),թե կմերժվի,այսինքն կընդունվի որպես օտար (ոչ հիստոհամատեղելի): Հյուսվածքների մերժումը դոնոր բջիջների մակերեսին գտնվող սպիտակուցների հանդեպ առաջացած իմուն պատասխանի արդյունք է (այդ սպիտակուցները կոչվում են հիստոհամատեղելիության հակածիններ` մկների H-անտիգեններ): Նրանք մերժման համար գլխավոր պատասխանատու գեներն են, չնայած կան բազմաթիվ այլգեներ, այդ պատճառով համակարգը կոչվում է գլխավոր հիստոհամատեղելիության համակարգ (MHC): Նման հակածիններ հայտնաբերվել են մարդու լեյկոցիտների վրա, այստեղից էլ այդ համակարգի երկրորդ անունը`Human Leukocyte Antigens(HLA):
Հիստոհամատեղելիությունը MHC-ն գեների մի խումբ է,որի արգասիքները կարևոր դեր են խաղում իմուն պատասխանի կայացման և սեփականը օտարից տարբերելու գործում: Կոչվում են հիստոհամատեղելիության գեներ,քանի որ նրանցով է որոշվում,թե փոխպատվաստված հյուսվածքը կընդունվի տիրոջ կողմից (հիստոհամատեղելի),թե կմերժվի,այսինքն կընդունվի որպես օտար (ոչ հիստոհամատեղելի): Հյուսվածքների մերժումը դոնոր բջիջների մակերեսին գտնվող սպիտակուցների հանդեպ առաջացած իմուն պատասխանի արդյունք է (այդ սպիտակուցները կոչվում են հիստոհամատեղելիության հակածիններ` մկների H-անտիգեններ): Նրանք մերժման համար գլխավոր պատասխանատու գեներն են, չնայած կան բազմաթիվ այլգեներ, այդ պատճառով համակարգը կոչվում է գլխավոր հիստոհամատեղելիության համակարգ (MHC): Նման հակածիններ հայտնաբերվել են մարդու լեյկոցիտների վրա, այստեղից էլ այդ համակարգի երկրորդ անունը`Human Leukocyte Antigens(HLA):
MHC-ի կարևորագույն դերը ձեռք բերովի իմուն պատասխանի ձևավորման մեջ է, քանի որ Th և Tc բջիջները չեն կարող ճանաչել օտար հակածինը,երբ այն մենակ է,այլ ճանաչում են միայն MHC մոլեկուլների հետ կապակցված վիճակում: Այսպիսով, եթե հակածին- MHC համակցությունը տեղի չունենա, ապա ձեռքբերովի իմուն պատասխան չի զարգանա:
MHC մոլեկուլները օժտված են բարձր պոլիմորֆիզմով, և տարբեր անհատների մոտ էքսպրեսիայի են ենթարկվում (դրսևորվում են) տարբեր ալելներ:Ուստի այդպիսով նրանք բնորոշում են հակածինների այն տեսականին, որոնց հանդեպ անհատը կարող է առաջացնել իմուն պատասխան:Այսպիսով նրանք ազդում են ինֆեկցիոն հիվանդացության վրա:Այլ կերպ ասած`նրանք որոշում են անհատի զգայունությունը հիվանդությունների նկատմամբ:
MHC գեների տեղակայումը
MHC կամ HLA գեները մարդու մոտ տեղակայված են 6-րդ քրոմոսոմի վրա երեք ռեգիոններում`MHC I,II և III ռեգիոններում:Յուրաքանչյուր ռեգիոն կարող է պարունակել գեների բազմաթիվ լոկուսներ: Յուրաքանչյուր ռեգիոնի գեներ առանձին դասի HLA մոլեկուլներ են կոդավորում:
MHC I դասի գեներ
Սրանք կոդավորում են գլիկոպրոտեինների սինթեզ,որոնք էքսպրեսիայի են ենթարկվում բոլոր կորիզավոր բջիջներում: Առաջին դասի մոլեկուլների գլխավոր ֆունկցիան պեպտիդային հակածինները Tc-ին (ցիտոտոքսիկ բջիջներ) ներկայացնելն է:
MHC II դասի գեներ
Սրանք կոդավորում են գլիկոպրոտեինների սինթեզ, որոնք էքսպեսիայի են ենթարկվում հիմնականում հակածին ներկայացնող բջիջների վրա (մակրոֆագեր,դենդրիտային բջիջներ և B-լիմֆոցիտներ):Երկրորդ դասի մոլեկուլները ներկայացնում են հակածինը Th-բջիջներին:
MHC IIIդասի գեներ
Սրանք կոդավորում են մոլեկուլներ,որոնք իմունային պատասխանում օժանդակող նշանակություն ունեն. այդպիսիք են կոմպլեմենտի որոշ բաղադրիչները, բորբոքիչ մոլեկուլներ, TNF`ուռուցթի նեկրոզի գործոն, HSP`ջերմային շոկի սպիտակուցներ և այլն: I ռեգիոնը ամենամեծն է, այստեղ գեները դասավորված են երեք հիմնական լոկուսներում`A,B,C: II ռեգիոնը բաժանվում է DP,DQ և DR լոկուսների:I և II դասի սպիտակուցներն ունեն կառուցվածքային նմանություն, երկուսն էլ կապված են բջջաթաղանթին,երկուսն էլ ունեն անմիջական մասնակցություն իմունային պատասխանի մեջ:III դասի մոլեկուլներն ունեն օժանդակող նշանակություն իմունային պատասխանում և արտադրվում են լուծելի տեսքով: I և II ռեգիոնների A,B,C և DP,DQ,DR գեների արգասիքները կոչվում են "դասական" MHC մոլեկուլներ, քանի որ ունեն անմիջական մասնակցություն իմուն պատասխանի կայացման մեջ: "Դասական" գեներին ի լրումն հայտնաբերվել են "ոչ դասական" գեներ, որոնք գտնվում են առաջին դասի լոկուսի B և C գեների միջև:Դրանք HLA-E,F,G գեներն են:E գենով կոդավորվող մոլեկուլները էքսպրեսիայի են ենթարկվում հասուն,հանգստի վիճակում գտնվող T-լիմֆոցիտների և մարդու կարցինոմայի բջիջների վրա: F գենով կոդավորվող հակածինները դեռևս հայտնաբերված չեն:G գեները էքսպրեսիայի են ենթարկվում խորիոկարցինոմայի բջիջների վրա,խրոնիկ ցիտոտրոֆոբլաստոզի ժամանակ բջիջների վրա, իսկ ֆիզիոլոգիական նշանակությունը կապված է վերարտադրողության հետ.այն էքսպրեսիայի ենթարկվելով միայն ընկերքի տրոֆոբլաստի բջիջների կողմից` NK-բջիջների ինհիբիտոր է հանդիսանում, որը ընկերքին կայունություն է տալիս NK-բջիջների հանդեպ: E և G գեների սպիտակուցները կապվում են պեպտիդ-հակածինների հետ,ընդ որում այդ համալիրները ճանաչվում են NK-բջիջների կողմից:HLA-E-ի ճանաչումը կատարվում է CD94 ընկալիչի միջոցով: Այս "ոչ դասական" գեների պոլիմորֆիզմի հարցը դեռ սպասում է պատասխանի,ինչպես նաև նրանց կենսաբանական նշանակության հարցը:
MHC գեների ժառանգումը
MHC լոկուսները օժտված են բարձր պոլիմորֆիզմով, այսինքն, պոպուլացիայում յուրաքանչյուր լոկուսի համար գոյություն ունեն բազմաթիվ այլընտրանքային տարբերակներ: Տվյալ գենը անհատի մոտ կարող է լինել հոմոզիգոտ (երկու քրոմոսոմների վրա գտնվում է տվյալ գենի նույն ալելը) կամ հետերոզիգոտ (երկու քրոմոսոմների վրա գտնվում է տվյալ գենի տարբեր ալելներ): Պոպուլացիայում անհատների մեծ մասը հետերոզիգոտ են ըստ MHC գեների: Նույն լոկուսում հայտնաբերված տարբեր ալելները համարակալվում են թվանշաններով, օր.`մի որևէ քրոմոսոմի վրա կարող են լինել հետևյալ գեները`MHC-A2, MHC-B5, MHC-C32, MHC-DR3, MHC-DP1, MHC-DQ3: Մյուս քրոմոսոմի վրա(զույգի վրա) կարող է լինել նման կամ տարբեր կազմ:
Մեկ քրոմոսոմի վրա գտնվող MHC ալելների ամբողջությունը, որն իր մեջ ներառում է բոլոր լոկուսները,կոչվում է հապլոտիպ: Յուրաքանչյուր ծնողից ժառանգվում է մեկական հապլոտիպ: Արդյունքում առաջացած դիպլոիդ կազմը սովորաբար հետերոզիգոտ է լինում:
Մեկ ուրիշ առանձնահատկություն. Հապլոտիպի տարբեր լոկուսները բավականին մոտ են տեղադրված իրար, ուստի բեղմնավորման ժամանակ գենետիկական ռեկոմբինացիայի հավանականությունը ընդամենը 0.5% է: Դա նշանակում է,որ ծնողի մեկ քրոմոսոմի հապլոտիպը ամբողջությամբ է ժառանգվում, որպես այդպիսին:
Գենետիկ էքսպրեսիայի տարբեր ձևեր կան, այդ թվում աուտոսոմ ռեցեսիվ և դոմինանտ ձևերը: Սակայն այս գեների դեպքում ժառանգումը կատարվում է կոդոմինանտ սկզբունքով, այսինքն նույն բջջի մեջ դրսևորվում են և մայրական և հայրական հապլոտիպները:Սիբլինգների միջև ընդամենը 25% հավանականություն կա, որ նրանք կլինեն հիստոհամատեղելի, և, իհարկե, երեխաներից ոչ մեկը չի կարող հիստոհամատեղելի լինել ծնողի հետ:
MHC մոլեկուլների կառուցվածքը
I դասի մոլեկուլներ
Բոլոր MHC I դասի մոլեկուլները գլիկոպրոտեիններ են և կազմված են մեկ մեծ α-շղթայից, որը կապակցված է ոչ MHC դասի մոլեկուլի` β2-միկրոգլոբուլինի հետ. վերջինս կոդավորված է 15-րդ քրոմոսոմի վրա գտնվող գենով:
α-շղթան տրանսմեմբրանային գլիկոպրոտեին է, որը կոդավորվում է HLA-A,B և C գեներով: β2-միկրոգլոբուլինի հետ կապն անհրաժեշտ է թաղանթի վրա էքսպրեսիայի համար: α-շղթան պարուրված է և ունի երեք արտաբջջային դոմեններ`α1, α2 և α3, յուրաքանչյուրը մոտ 90 ամինաթթվային հաջորդականությամբ:Անյուհետև նրանց հաջորդում է տրանսմեմբրանային դոմենը`46 հիդրոֆոբ ամինաթթվային կազմով և ցիտոպլազմատիկ կարբօքսիլ-ծայրային հատվածը դոմենը` 30 ամինաթթվային կազմով: β2-միկրոգլոբուլինն ունի միայն արտաբջջային դոմեն, որը նման է α3 դոմենին և նրա հետ փոխազդում է ուժեղ ոչ կովալենտ կապով:
α1 և α2 դոմենների միջև ձևավորվում է ճեղք, որը հակածին կապող հատված է: Այդ ճեղքի(Բյորկմանի ճեղք) հատակը կազմված է 8 հակազուգահեռ β-ծալքերից, իսկ պատերը կազմված են α-պարույրից:Այս ճեղքում կարող է տեղավորվել 8-10 ամինաթթվային պեպտիդ: α3 դոմենը և β2-միկրոգլոբուլինն իրար հետ կազմավորում են իմունագլոբուլինի նման կառուցվածք, որոնցից յուրաքանչյուրը բաղկացած է երկու β-ծալքերից: Այդ երկու դոմենների և իմունոգլոբուլինների կոնստանտ հատվածների միջև կա զգալի հոմոլոգիա:Ընդհանուր առմամբ I դասի մոլեկուլի կառուցվածքը նման է Ig մոլեկուլի կառուցվածքի: α1,-α2-ը նման են հակածին կապող հատվածին, իսկ α3 դոմենը և β2-միկրոգլոբուլինը նման են կոնստանտ հատվածին: Իմունոգլոբուլինի նման` այս մոլեկուլը ևս քայքայվում է պապաին ֆերմենտով` տրանսմեմբրանային հատվածից քիչ պրոքսիմալ հատվածում: Քանի որ I դասի մոլեկուլները նման են Ig-ի, ուստի MHC I դասի մոլեկուլները և β2-միկրոգլոբուլինը համարվում են Ig սուպերընտանիքի անդամ:
α3 դոմենը մասնակցում է Tc բջջի CD8 ընկալիչի հետ կապ հաստատմանը: β2-միկրոգլոբուլինի բացակայության պայմաններում I դասի MHC մոլեկուլները էքսպրեսիայի չեն ենթարկվում բջջի թաղանթի վրա:
II դասի մոլեկուլներ
MHC II դասի մոլեկուլների կառուցվածքը բավականին նման է I դասի մոլեկուլներին: Այնուամենայնիվ, այն հետերոդիմեր է` կազմված α և β-շղթաներից: Այն նույնպես ունի արտաբջջային ծալվածք,որը նման է իմունոգլոբուլինի`հակածին կապող հատվածի, այդ իսկ պատճառով այն ևս համարվում է Ig սուպերընտանիքի անդամ:
α և β-շղթաները տրանսմեմբրանային գլիկոպրոտեիններ են, յուրաքանչյուրը երկուական արտաբջջային դոմեններով`համապատասխանաբար α1 ,α2 և β1, β2:Այնուհետև յուրաքանչյուր շղթան ունի տրանսմեմբրանային և ցիտոպլազմատիկ հատված: α1 և β1-ի փոխազդեցությունը նման է I դասի մոլեկուլի α1 և α2 դոմենների փոխազդեցությանը և ստեղծում են Բյորկմանի ճեղքի α-պարույրը: α2 և β2-ի կապը նման է I դասի մոլեկուլի α3 դոմենի և β2-միկրոգլոբուլինի փոխազդեցությանը:MHC II-ի պեպտիդ կապող հատվածը MHCI-ի նույնանման հատվածին, բայց լայն է և երկու ծայրից բաց, ուստի կարող է կապել ավելի մեծ թվով ամինաթթվային հաջորդականությամբ պեպտիդ:
MHC մոլեկուլների պոլիմորֆիզմը 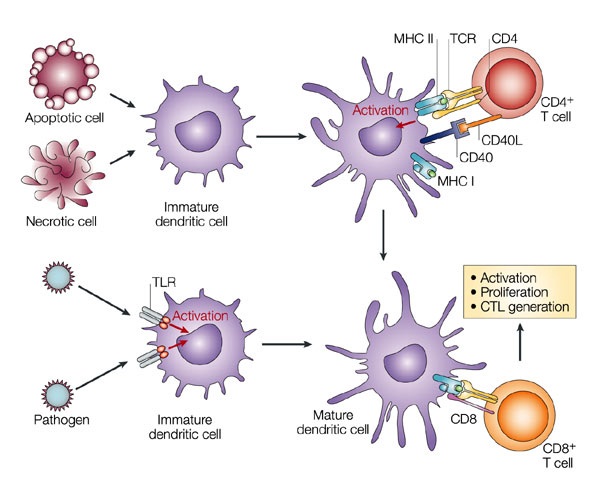
MHC մոլեկուլները փոխազդում են հակածնային պեպտիդի հետ և ներկայացնում այն T-բջջին: Քանի որ պեպտիդները շատ բազմազան են` ստացված ամենատարբեր հակածիններից, ուստի անհրաժեշտ է, որպեսզի այս մոլեկուլները ևս լինեն բազմազան`այդ ամենը ներկայացնել կարողանալու համար: Դա ի շնորհիվ MHC գենոմի հատկության է, ըստ որի այն կարող է էքսպրեսիայի ենթարկել MHC մոլեկուլների լայն ընտրանի` ամենաբազմազան հակածնային սպեցիֆիկությամբ:
MHC մոլեկուլների պոլիմորֆիզմի պատճառները երեքն են`
1.Բազմաթիվ գեների լոկուսներ-և I և II դասի մոլեկուլները կոդավորվում են առանձին տեղակայված բազմաթիվ գեներով: Յուրաքանչյուր լոկուսի գեներ կոդավորում են նման, բայց ոչ նույնական (իդենտիկ) շղթաներ:
2.Տարբեր լոկուսների բազմաթիվ ալելներ-նույն լոկուսի գեների համար գոյություն ունեն բազմաթիվ ալելներ. այդ ալելներն իրարից տարբերվում են իրենց ԴՆԹ-ի հաջորդականության 5-10%-ով: Տարբեր ալելները հանդիպում են տարբեր անհատներ մոտ և կոդավորում տարբեր կառուցվածքով MHC մոլեկուլներ,որոնք ունեն տարբեր հակածնային սպեցիֆիկություն: Օր.` մարդու I դասի ռեգիոնում հայտնի է մոտ 60-A ալելներ, 110-B ալելներ և 40-C ալելներ: β2-միկրոգլոբուլինը պոլիմորֆ չէ:
II դասի մոլեկուլները օժտված են բարձր աստիճանի պոլիմորֆիզմով: Օր.` DR լոկուսի β-շղթայի գեների թիվը կարող է 2-ից 5-ը լինել տարբեր հապլոտիպերում: Հայտնի են DR լոկուսի β-շղթայի 122 ալելներ: Այնուամենայնիվ տվյալ տեսակի մոտ DR լոկուսի α-շղթան նույնական է բոլորի մոտ:
3.Կոդոմինանտ էքսպրեսիա-MHC գեների լոկուսները էքսպրեսիայի են ենթարկվում կոդոմինանտ ձևով, այսինքն` երկու հապլոտիպերը, որոնք պտուղը ստանում է` մեկը հորից, մյուսը` մորից, դրսևորվում են հավասարապես բոլոր բջիջների վրա: Այսպիսով պտուղը էքսպրեսիայի է ենթարկում 6 I դասի α-շղթա և 12 II դասի α/β շղթաներ: Այդ բոլորը դրսևորվում են միևնույն բջջի մակերեսին:
II դասի մի լոկուսի սահմանում մայրական α-շղթան կարող է կապվել հայրական β-շղթայի հետ և հակառակը, ուստի դա յուրաքանչյուր լոկուսում 4 տարբերակ ունենալու հնարավորություն է տալիս: Այս ամենը հնարավորություն է տալիս ունենալ MHC մեծ պոլիմորֆիզմ. մկների մետ MHC տեսականին 1012 է, մարդու մոտ ավելի շատ:
MHC պոլիմորֆիզմի կարևորությունը
MHC գեների հաջորդականության ուսումնասիրությունը ցույց է տվել, որ ամբողջ շղթան պոլիմորֆ չէ, այլ հատկապես պոլիմորֆիզմը բնորոշ է հակածնի պեպտիդը կապող դոմեններին` α1 և α2 դոմեններին I դասի համար և α1 և β1-դոմեններին II դասի համար: Ուստի նպատակը բազմազան սպեցիֆիկությամբ հակածին կապող հատվածների առկայությունն է:
MHC մոլեկուլների կողմից հակածին կապելը
Չնայած MHC գեների հարյուրավոր ալելների առկայությանը, մեկ անհատը կարող է միջինում էքսպրեսիայի ենթարկել 6 տիպի I դասի մոլեկուլ և 12 տիպի II դասի մոլեկուլ: Բայց նույնիսկ այս սահմանափակ քանակը կարող է զգալի թվով պեպտիդների հետ փոխազդել, քանի որ հակածին կապելու համար անհրաժեշտ է յուրահատուկ ամինաթթվային հաջորդականություն, որը կարող է շատ պեպտիդներում առկա լինել: Այսպիսով, մեկ MHC մոլեկուլը կարող է կապել բազմաթիվ պեպտիդներ, պայմանով, որ այդ պեպտիդների ամինաթթվային հաջորդականությունը նույնն է մոլեկուլի որոշակի նույն դիրքում, չնայած մյուս հատվածների ամինաթթվային հաջորդականությունը կարող է տարբեր լինել:
Հակածնի կապվելը MHC I դասի մոլեկուլների հետ
I դասի մոլեկուլները կապվում են էնդոգեն (մարդու սեփական բջիջներում որոշակի իրավիճակներում արտադրված պեպտիդներ) պեպտիդների հետ: Օր.` վիրուսային վարակը բերում է վիրուսի գենոմի ներմուծմանը տիրոջ գենոմ: Ապա վիրուսային գենոմն օգտագործում է տիրոջ մեխանիզմները` սեփական սպիտակուցների սինթեզի համար: Սրանք ակնհայտորեն օտար սպիտակուցներ են: Նման իրավիճակ է նաև քաղցկեղային բջիջների կամ այլ` գենետիկորեն փոփոխված բջիջների դեպքում: Այս պեպտիդները մարսվում են ցիտոզոլում, հարմար պեպտիդային հատվածը տեղափոխվում է էնդոպլազմատիկ ցանց, որտեղ միանում է I դասի մոլեկուլի հետ: Այս համալիրը այնուհետև գնում է դեպի բջջի մակերես` էքսպեսիայի է ենթարկվում բջջաթաղանթի վրա:
MHC I դասի մոլեկուլների բջջային բաշխումը
MHC I դասի մոլեկուլները էքսպրեսիայի են ենթարկվում բոլոր կորիզավոր բջիջների վրա, սակայն էքսպրեսիայի աստիճանը տարբեր է: Նրանք առավելագույն մակարդակով դրսևորվում են լիմֆոցիտների վրա, իսկ լյարդի բջիջների,նեյրոնների և միոցիտների վրա` նվազագույն մակարդակով: Ուստի լյարդի փոխպատվաստումը հազվադեպ է մերժվում:
Յուրաքանչյուր բջիջ էքսպրեսիայի է ենթարկում յուրաքանչյուր MHC I մոլեկուլի 105 կրկնօրինակներ: Հետերոզիգոտ անձը կունենա 6 տարբեր տեսակի MHC I մոլեկուլներ` ի շնորհիվ կոդոմինանտ էքսպրեսիայի: Եվ յուրաքանչյուր մոլեկուլի տեսակը, իրենց ալելային տարբերության շնորհիվ, կարող է ճանաչել տարբեր բազմազան պեպտիդներ: Այսպիսով մի բջիջը, ունենալով իր մակերեսին MHC I դասի մոլեկուլների այսպիսի մեծ քանակ, կարող է միաժամանակ ներկայացնել իր մակերեսին բազմաթիվ պեպտիդներ: Սրանք Ճանաչվում են Tc լիմֆոցիտների կողմից և ձևավորում բջջային իմուն պատասխան: Փաստերը վկայում են, որ մոտ 100 հակածնի պեպտիդ-MHC I մոլեկուլ համալիրի առկայությունը բավարար է բջջի ճանաչման և լիզիսի համար:
Հակածնի պեպտիդի և MHC I դասի մոլեկուլների փոխազդեցությունը
Հակածին կապող հատվածը ճեղքի նման է, որը փակ է երկու կողմից: Սա սահմանափակում է (ռեստրիկցիայի է ենթարկում) կապվող պեպտիդի երկարությունը մինչև 8-10 ամինաթթվային հաջորդականության (իդեալական է նոնամերը): Բացի այդ 8-10 ամինաթթուներից` պեպտիդները պետք է ունենան հիդրոֆոբ մնացորդներ C-ծայրում և N-ծայրից 2-րդ կամ 2-րդ և 3-րդ դիրքերում: Այս մնացորդների (սովորաբար իզոլեյցին, լեյցին կամ վալին) կողմնային շղթաները ներմուծվում են MHC I ձեղքի մեջ` այդպիսով խարսխելով պեպտիդը ակոսի մեջ: Այդ պատճառով այդ ծայրային մնացորդները կոչվում են խարսխող մնացորդներ: Երբեմն գլիցինը, պրոլինը կամ թիրոզինը, լինելով N-ծայրին մոտ դիրքերում, ևս կարող են խաղալ խարսխող մնացորդի դեր: Ընդհանրապես ցանկացած պեպտիդ,որն ունի ճիշտ երկարություն և խարսխող մնացորդներ,կարող է կապվել MHC I դասի մոլեկուլին:C-ծայրային մնացորդները սովորաբար հաստատուն են այն պեպտիդներում, որոնք կապվում են MHC I դասի մոլեկուլների հետ:
Կապված պեպտիդը խարսխված է N- և C-ծայրերով ճեղքի մեջ, իսկ մեջտեղի հատվածը նկատելի կոնտակտ չունի MHC մոլեկուլի հետ, ուստի անգամ մի փոքր մեծ պեպտիդը կարող է տեղավորվել այնտեղ`ճիշտ խարսխող ծայրային մնացորդների դեպքում, քանի որ ընդհանուր պեպտիդը դուրս է գալիս ճեղքի սահմանից: Ինչպես նաև նույնանման խարսխող ծայրային մնացորդներով, բայց տարբեր միջին հատվածով պեպտիդները կարող են կապվել նույն MHC մոլեկուլի հետ: Հենց այս մեջտեղի ամինաթթուներն էլ, ազատ լինելով, կարող են անմիջապես փոխազդել TCR-ի (T-cell receptor)` T-բջջային ընկալիչի հետ:
Հակածնի կապվելը MHC II դասի մոլեկուլների հետ
II դասի մոլեկուլները կապում են էկզոգեն պեպտիդները, այսինքն հակածինը, որը մուտք է գործում դրսից, ինչպիսիք են բակտերիաները, սնկերը և պարազիտները կամ նրանք արգասիքները: Այս հակածինները քայքայվում են ներբջջային պրոցեսինգի (մարսողություն) արդյունքում և հարմար պեպտիդային հատվածը կապվում է MHC II մոլեկուլի հետ: Սովորաբար այդ պեպտիդային հատվածները պատկանում են մանրէների թաղանթային սպիտակուցներին, որոնք ինտերնալիզացիայի են ենթարկվում (մտնում են բջջի մեջ) էնդոցիտոզի կամ ֆագոցիտոզի միջոցով:
MHC II դասի մոլեկուլների բջջային բաշխումը
Այս դասի մոլեկուլները հիմնականում գտնվում են հակածին ներկայացնող բջիջների վրա, այսինքն`մակրոֆագերի, B-բջիջների և դենդրիտային բջիջների վրա: Այս բջիջներից որոշը էքսպրեսիայի են ենթարկում II դասի մոլեկուլները կոնստիտուտիվ կերպով:
MHC II դասի մոլեկուլները ներկայացնում են իրենց կապված էկզոգեն պեպտիդը Th-բջիջներին, որոնք ակտիվանալով օգնում են և B- և Tc-բջիջներին զարգացնելու հումորալ կամ բջջային իմուն պատասխան:
MHC II դասի մոլեկուլների պեպտիդ կապող ճեղքը ավելի մեծ է և երկու կողմից բաց: Ուստի այն կարող է կապել 13-18 ամինաթթվային մնացորդներ:Այստեղ չկան խարսխող մնացորդներ, փոխարենը ամբողջ պեպտիդի երկայնքով գործում են ջրածնային կապեր: Կենտրոնական 7-10 ամինաթթուները սովորաբար ամենակարևորն են և հարուստ են արոմատիկ ու հիդրոֆոբ ամինաթթուներով:
MHC էքսպրեսիայի կարգավորումը
MHC I և II դասի էքսպրեսիան տարբեր է և էքսպրեսիայի աստիճանով և բջջային կազմով: Դա MHC էքսպրեսիայի կարգավորման շնորհիվ է:
Եվ I և II դասի գեները իրենց 5’-ծայրային մասում ունեն պրոմոտորներ, որոնք կարգավորվում են դրական և բացասական կարգավորիչների` տրանսկրիպցիայի գործոնների միջոցով: Երկու այսպիսի գործոններ հայտնաբերված են. դրանք են CII TA և RF-X` երկուսն էլ կապվում են MHC II գեների պրոմոտորներին և ուժեղացնում նրանց տրանսկրիպցիան: Այս գործոնների պակասուրդը բերում է T-բջիջների առաջացման, որոնք իրենց մակերեսին չունեն MHC II մոլեկուլներ: Այսպիսի պացիենտները տառապում են ծանր իմունային անբավարարությամբ: Այս համախտանիշը կոչվում է "մերկ լիմֆոցիտների համախտանիշ":
Ցիտոկինները ևս ազդում են MHC էքսպրեսիայի վրա: IF-g-ն ուժեղացնում է I և II դասի մոլկուլների էքսպրեսիան` խթանելով CII TA գործոնը և մեկ այլ յուրահատուկ տրանսկրիպցիայի գործոն` I դասի գեների պրոմոտորի համար: IL-4-ը խթանում է հանգստացող B-բջիջներում II դասի մոլեկուլների էքպրեսիան:
Կորտիկոստերոիդները և պրոստագլանդինները ընկճում են II դասի մոլեկուլների էքսպրեսիան: Շատ վիրուսային վարակներ ընկճում են I դասի մոլեկուլների էքսպրեսիան` խանգարելով նրանց հյուսվածքների էքսպրեսիային և/կամ հավաքման մեխանիզմին: Ցիտոմեգալովիրուսային վարակի դեպքում վիրուսի սպիտակուցները կապվում են β2-միկրոգլոբուլինին` արգելակելով MHC I էքսպրեսիան: Ադենովիրուսները ընկճում են TAP-1 TAP-2 գեների տրանսկրիպցիան` բերելով նրանց ցածր արտադրության, որը նույնպես իջեցնում է MHC I էքսպրեսիան: Վիրուսի համար նպատակահարմար է ընկճել MHC I դասի էքսպրեսիան, քանի որ այդ դեպքում այն կխուսափի MHC I դասի միջոցով միջնորդված իմուն պատասխանից:
MHC-ն և հիվանդությունների նկատմամբ ընկալունակությունը
Շատ HLA ալելներ կապակցված են որոշ հիվանդություններով` հատկապես աուտոիմունային հիվանդություններով հիվանդանալու բարձր ընկալունակության հետ:
Հիվանդանալու հարաբերական ռիսկը տարբեր է.օրինակ` HLA B-27 ունեցողների մոտ անկիլոզացնող սպոնդիլոարթրիտով հիվանդանալու ռիսկը 50 անգամ մեծ է, քան չունեցողների մոտ: Այս ալելի էքսպրեսիան ունի կարևոր, բայց ոչ բացառիկ դեր.գենետիկ և միջավայրի գործոնները ունեն հավասար նշանակություն:
Հիվանդություններով հիվանդանալու համար MHC-կապակցվածությունը կարող է երկու հնարավոր պատճառ ունենալ`
1.Յուրահատուկ MHC-ալելը կարող է որոշակի ախտածին հարուցիչի նկատմամբ իմուն պատասխան ձևավորելու գործում ներգրավված լինել: Եթե պաթոգենի գլխավոր հակածնային պեպտիդը նմանակում է սեփական MHC մոլեկուլին, ապա T-բջիջը համապատասխան ռեակցիա չի տա նրան, քանի որ տոլերանտ է դառնում նրա նկատմամբ:
2.Տեսակի մեջ MHC պոլիմորֆիզմի պակասումը կարող է տվյալ տեսակի` ինֆեկցիոն հիվանդություններով հիվանդանալու նախատրամադրվածության պատճառ դառնալ:
Էլեկտրոնային նյութի սկզբնաղբյուրը ՝ Doctors.am
Նյութի էլէկտրոնային տարբերակի իրավունքը պատկանում է Doctors.am կայքին



