






 Արյունաստեղծումը (հեմոպոեզ) անընդհատ պրոցես է, որն սկսվում է սաղմնային զարգացման վաղ փուլերում և շարունակվում մինչև մահ։ Ստեղծված երիտասարդ բջիջները փոխարինում են ծերացած և անկենսունակ արյան բջիջներին, որոնց կյանքի տևողությունը (բացառությամբ հիշողության լիմֆոցիտների) անհամեմատ կարճ է։ Արյունաստեղծման կառուցողական և էներգիական փոխանակության համար անհրաժեշտ են սպիտակուցների, ճարպերի, ածխաջրերի, նուկլեոտիդների, վիտամինների և հանքային աղերի, մասնավորապես միկրոտարրերի անընդհատ ներհոս։ Վիտամիններն ու միկրոտարրերը ներգրավվում են նյութափոխանակության որոշակի օղակների մեջ, առանց որի արյունաստեղծման պրոցեսը տվյալ փուլում խափանվում է, առաջ բերելով արյան բջիջների հետագա զարգացման ու հասունացման լուրջ խանգարումներ։
Արյունաստեղծումը (հեմոպոեզ) անընդհատ պրոցես է, որն սկսվում է սաղմնային զարգացման վաղ փուլերում և շարունակվում մինչև մահ։ Ստեղծված երիտասարդ բջիջները փոխարինում են ծերացած և անկենսունակ արյան բջիջներին, որոնց կյանքի տևողությունը (բացառությամբ հիշողության լիմֆոցիտների) անհամեմատ կարճ է։ Արյունաստեղծման կառուցողական և էներգիական փոխանակության համար անհրաժեշտ են սպիտակուցների, ճարպերի, ածխաջրերի, նուկլեոտիդների, վիտամինների և հանքային աղերի, մասնավորապես միկրոտարրերի անընդհատ ներհոս։ Վիտամիններն ու միկրոտարրերը ներգրավվում են նյութափոխանակության որոշակի օղակների մեջ, առանց որի արյունաստեղծման պրոցեսը տվյալ փուլում խափանվում է, առաջ բերելով արյան բջիջների հետագա զարգացման ու հասունացման լուրջ խանգարումներ։
Վիտամին В12-ը կոչվում է նաև Կասլի արտաքին գործոն։ Այն, ընդունած սննդի հետ ստամոքս մտնելով, միանում է ստամոքսի լորձաթաղանթի կողմից արտադրվող Կասլի ներքին գործոն կոչվող գաստրոմուկոպրոտեիդների հետ և դրանով վիտամինը պաշտպանում մարսողական ֆերմենտների ազդեցությունից և մանրէների կողմից յուրացումից։ Աղիքներից վիտամին В12-ը ներծծվում է լյարդի մեջ և կուտակվում լյարդում։ Լյարդից աստիճանաբար փոխադրվում է ոսկրածուծ և ապահովում բաժանվող բջիջների նուկլեինաթթուների սինթեզը։ Հաշվված է, որ մեկ բջջի միտոզի համար անհրաժեշտ է մեկ մոլեկուլ վիտամին B12: Նրա անբավարարությունն առաջին հերթին, որպես ամենաարագ կիսվող բջիջների, արտահայտվում է էրիթրոիդ շարքի բջիջների վրա, առաջ բերելով վիտամին В12- անբավարարության հիպերքրոմ սակավարյունություն։
Վիտամին В9 կամ ֆոլաթթուն մասնակցում է արյունաստեղծ բջիջների ԴՆԹ-ի սինթեզին։ Նրա անբավարարությունը դարձյալ դրսևորվում է առաջին հերթին էրիթրոպոեզի բնականոն ընթացքի շեղումով, որի հետևանքով էրիթրոցիտների կյանքի տևողությունը խիստ կրճատվում է։
Վիտամին В6-ը որպես կոֆերմենտ մասնակցում է հեմի սինթեզին, ուստի նրա անբավարարությունն ուղեկցվում է հիպոքրոմ սակավարյունությամբ։
Վիտամին B2-ն ապահովում է էրիթրոպոեզի բնականոն ընթացքը, կարգավորելով արյունաստեղծ բջիջների օքսիդավերականգնման ռեակցիաները։ Անբավարարության դեպքում զարգանում է հիպոռեգեներատոր սակավարյունություն։
Վիտամին C-ն արագացնում է աղիքներից երկաթի ներծծումը, իսկ ոսկրածուծում` նպաստում հեմի սինթեզին։ ֆոլաթթ վի հետ համագործակցելով, վիտամին C-ն արագացնում է նաև միտոզի պրոցեսը ոսկրածուծում։
Վիտամին E-ն որպես հակաօքսիդանտ, պահպանում է էրիթրոցիտների լիպիդային թաղանթների ամբողջականությունը։
Վիտամին PP-ն մասնակցում է թաղանթների ֆոսֆոլիպիդների սինթեզին և որպես ջրածնի փոխադրիչ, կարևոր դեր ունի նաև արյունաստեղծ բջիջների շնչառության պրոցեսում։ Հեմոպոեզի բնականոն ընթացքի համար կարեոր նշանակություն ունեն նաև մի շարք միկրոտարրեր պղինձը, նիկելը, կոբալտը, սելենը, ցինկը և այլն։
Պղնձի ներկայությամբ երկաթե արագությամբ ներծծվում է աղիքներից և կուտակվում լյարդում և ռետկուլյար բջիջներում, իսկ հեմի սինթեզի պրոցեսում պղինձը նպաստում է նաև երկաթի յուրացմանը։
Նիկելը և կոբալտը նույնպես մասնակցում են հեմոգլոբինի սինթեզին։ Այն վայրերում, որտեղ հողն աղքատ է այս միկրոտարրերով, նկատվում է կենդանիների և մարդկանց սակավարյունություն։
Սելենը համագործակցում է վիտամին E-ի հետ՝ պահպանելով էրիթրոցիտների լիպիդային թաղանթները։
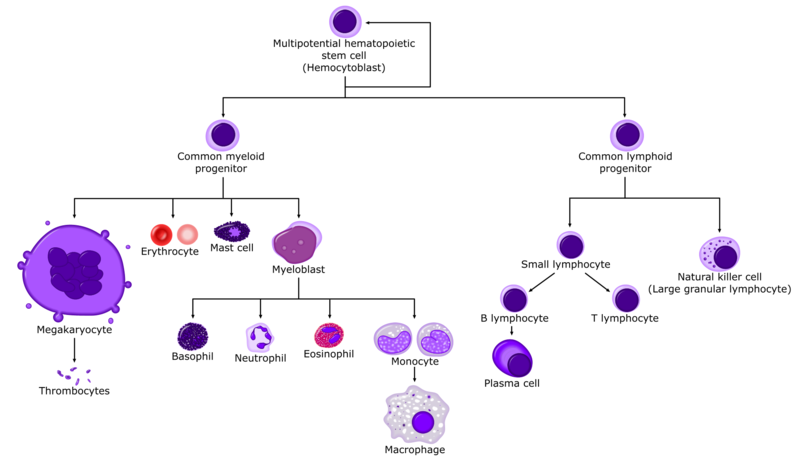
Ցինկը մտնում է էրիթրոցիտների կարբոանհիդրազա ֆերմենտի բաղադրության մեջ։ Օրգանիզմի ցինկի բաղադրության 75%-ը կապված է այդ ֆերմենտի հետ, որը մասնակցում է ածխաթթու գազի փոխադրմանը։ Ցինկը մասնակցում է նաև լեյկոպոեզի բնականոն ընթացքի կարգավորմանը։ Արյան բոլոր բջիջները ոսկրածուծում առաջանում են մեկ ընդհանուր մայր բջջից, որը կոչվում է ցողունային արյունաստեղծ բջիջ։ Ցողունային բջիջներն օժտված են ամբողջ կյանքի ընթացքում ինքնավերարտադրման և բազմացման չընդհատվող ունակությամբ, որի շնորհիվ առաջանում են բազմապոտենտ բջիջներ և տարբերակվում հեմոպոեզի համապատասխան ուղղություններով` 1) միելոպոեզի ուղղությամբ, որից սկսվում է էրիթրոպոեզի, գրանուլոպոեզի և մեգակարիոցիտոպոեզի նախնական տարբերակում, 2) T-լիմֆոցիտների նախորդ ձևերի տարբերակում, 3) B-լիմֆոցիտների նախորդ ձևերի տարբերակում։ Ըստ հյուսվածքային կուլտուրաներից ստացված տվյալների այս նախորդ ձևերը սկիզբ են տալիս առանձին միապոտենտ գաղութների, որոնցից յուրաքանչյուրում առանձին-առանձին ընթանում է էրիթրոպոեզը, գրանուլոպոեզի 3 ծիլերը, մոնոցիտոպոեզը, թրոմբոցիտոպոեզը, T- և B-լիմֆոցիտոպոեզը` իրենց հետագա զարգացման և հասունացման հաջորդական փուլերով։ T- և B-լիմֆոցիտների վերջնական տարբերակումը համապատասխանաբար տեղի է ունենում ուրցագեղձում և ավշային հանգույցներում։
Տարբերակվող բջիջներն իրենց թաղանթի վրա ձեռք են բերում սպիտակուցային բնույթի ընկալիչներ, որոնք զգայուն են իրենց համապատասխան հեմոպոետիկ հորմոնների, խթանիչների, արգելակիչների (ինհիբիտորներ), ինչպես նաև թիրեոտրոպ հորմոնի, տեստոստերոնի ածանցյալների, կատեխոլամինների և նեյրոմեդիատորների նկատմամբ։ Արյունաստեղծումն անընդհատ գտնվում է նյարդային և հումորալ կարգավորիչ մեխանիզմների հսկողության ներքո, որոնք, խթանելով կամ ընկճելով նրա ընթացքը, այն հարմարեցնում են օրգանիզմի առաջադրած պահանջներին։ Հումորալ ամենաէական կարգավորիչները յուրահատուկ հորմոններ են և ընդհանուր անունով կոչվում են հեմոպոետիններ։ Ըստ քիմիական բաղադրության և ֆունկցիայի հեմոպոետինները բաժանվում են երեք մեծ խմբի` էրիթրոպոետիններ, լեյկոպոետիններ և թրոմբոցիտոպոետիններ։ Դրանք արյան յուրաքանչյուր բջջի հետագա զարգացմանն ու տարբերակմանը համապատասխան ուղղություն են տալիս։ Հեմոպոետինները սինթեզվում են 5-րդ, 7-րդ և 17-րդ զույգ քրոմոսոմների համապատասխան գեների հսկողության տակ։ Այդ զեների գործունեության խանգարումն առաջ է բերում հեմոպոեզի տարբեր ծիլերի զարգացման ծանր խանգարումներ։
էրիթրոպոեզի կարգավորումը։ էրիթրոպոեզի հումորալ կարգավորման ամենահզոր գործոնը էրիթրոպոետին հորմոնն է, որն իր քիմիական կառուցվածքով գլիկոպրոտեիդ է։ Նրա 80-85%-ը սինթեզվում է երիկամների պերիտուբուլյար բջիջներում իսկ մնացածը լյարդի կուպֆերյան բջիջներում։ Այդ բջիջներում գոյություն ունի երկաթ պարունակող հատուկ սպիտակուց` հեմոպրոտեին, որը երկամի թթվածնով բավարար մատակարարման պայմաններում կապվում է 02 ի հետ և այդ միացության ձևով ճնշում էրիթրոպոետին արտադրող գենը։ Իսկ երբ երիկամի արյունամատակարարումը պակասում է, ապա հեմոպրոտեինից 02-ն անջատվում է և թթվածնազուրկ հեմոպրոտեինը ունենում է հակառակ ներգործություն՝ ակտիվացնում է էրիթրոպոետինի սինթեզն ապահովող գենը, որի գործունեությամբ էրիթրոպոետինի սինթեզը կարող է ուժեղանալ մինչև. 1000 անգամ։ էրիթրոպոետինը արագացնում է էրիթրոիդ շարքի նախորդ բջիջների բազմացումը և խթանում հեմոգլոբինի սինթեզը։ Սուր արյունահոսություններից հետո կամ բարձրադիր վայրերում ՝ էրիթրոպոեզը խթանվում է էրիթրոպոետինների սինթեզի ուժեղացման հաշվին։ էրիթրոպոետիններն ունեն իրենց արգելակիչները որոնք ճնշում են էրիթրոպոեզը։ Օրգանիզմի պահանջներին չհամապատասխանող էրիթրոցիտների քանակի ավելացումը խթանում է արգելակիչների սինթեզը, ինչպես դա լինում է, օրինակ, բարձրադիր վայրերից հարթավայրերն իջնելիս։ Արական սեռական հորմոնների ածանցյալները ուժեղացնում են էրիթրոպոետինների ներգործությունը, իսկ իգականները ընղհակառակը։. Դրանով է բացատրվում երկու սեռի էրիթրոցիտների քանակական տարբերությունը, որն սկսվում է սեռական հասունացումից հետո։ Կատեխոլամինները խթանում են էրիթրոիդ շարքի բջիջների բազմացումը։
Լեյկոպոեզի կարգավորումը։ Լեյկոպոեզը կարգավորվում է լեյկոպոետինների միջոցով, որոնց քիմիական բաղադրությունը և գոյացման վայրը դեռևս չի բացահայտված։ Գրանուլոցիտների և մոնոցիտների միապոտենտ գաղութները տարբերակվում են մակրոֆագերից, մոնոցիտներից և T-լիմֆոցիտներից անջատված գաղութախթանիչ գործոնների ազդեցության ներքո։ Մոնոցիտների և գրանուլոպոեզի 3 ճյուղերի` բազոֆիլների, էոզինոֆիլների և նեյտրոֆիլների գաղութներում լեյկոցիտների հետագա տարբերակումը շարունակվում է համապատասխանաբար մոնոցիտոպոետինների, բազոֆիլոպոետինների, էոզինոֆիլոպոետինների և նեյտրոֆիլոպոետինների մասնակցությամբ։ Լեյկոպոետինների սինթեզը խթանվում է ծերացած լեյկոցիտների և վնասված հյուսվածքների քայքայման արգասիքներով, նուկլեինաթթուներով, միկրոբներով և նրանց թույներով։ էոզինոֆիլոպոետինների ակտիվությունը մեծանում է նաև մակաբույծ որդերի արտադրած թույների ներկայությունից։
Հաստատված է, որ հասունացած նեյտրոֆիլներն արտադրում են լեյկոպոեզն ընկճող յուրահատուկ նյութեր` քեյլոններ, որոնք չափավորում են լեյկոպոեզի ուժեղացած ընթացքը։ Քեյլոնները ցածրամոլեկուլային միացություններ են և ճնշում են գրանուլոպոեզի նախորդ բջիջների ԴՆԹ-ի սինթեզը, հետևաբար, նաև նրանց բաժանման պրոցեսը։ Լեյկոզների բուժման խնդրում քեյլոնների գործնական կիրառումը մեծ հեռանկարներ է խոստանում։ Առողջ օրգանիզմում քեյլոնների և լեյկոպոետինների գործունեությունը գտնվում է որոշակի հավասարակշռության մեջ, որի շնորհիվ լեյկոպոեզը հարմարվում է օրգանիզմի պահանջներին։ Քեյլոններից բացի հասուն նեյտրոֆիլներն արտադրում են նաև մեկ այլ նյութ` լակտոֆերին, որը ճնշող ազդեցություն է ունենում նեյտրոֆիլների և մոնոցիտների տարբերակման վրա։ Մոնոցիտոպոեզի վրա դրդող ներգործություն ունեն կատեխոլամինները, իսկ Е խմբի պրոստագլանդինները և α ու β-ինտերֆերոնները արգելակող։
Թրոմբոցիտոպոեզի կարգավորումը։ Թրոմբոցիտոպոեզն սկսվում է մեգակարիոցիտային միապոտենտ գաղութներում, որոնք ձևավորվում են մեգակարիոցիտային գաղութային խթանիչ գործոնի մասնակցությամբ։ Այդ խթանիչը դրդում է թրոմբոցիտոպոեզի նախորդ բջիջների միտողը և տարբերակումը։ Առաջացած նոր բջիջների քանակն իր հերթին հակադարձ կապի սկզբունքով կարգավորում է խթանիչի արտադրությունը։ Ոսկրածուծում նրանց քանակը պակասելիս գաղութախթանիչի արտադրությունն ուժեղանում է և հատուցում այդ բջիջների պակասը։
Թրոմբոցիտոպոեզի հետագա բնականոն ընթացքը կարգավորվում է թրոմբոցիտոպոետինների ազդեցության ներքո, որոնց բաժանում են երկու խմբի` կարճատև և երկարատև ազդող թրոմբոցիտոպոետինների։ Երկարատև ազդող գործոնների ազդեցությունը տարածվում է թրոմբոցիտոպոեզի նախորդ բջիջների (պրոմեգակարիոբլաստ,մեգակարիոբլաստ և այլն) բազմացման և տարբերակման վրա, արագացնելով այդ պրոցեսները։ Իսկ կարճատև գործողները ապահովում են հասուն մեգակարիոցիտներից թրոմբոցիտների անջատման պրոցեսը։ Թրոմբոցիտոպոետինների սինթեզը դարձյալ հակադարձ կապի սկզբունքով կախված է արյան թրոմբոցիտների քանակից։ Երբ վերջինիս մակարդակը արյան 1մմ3 -ում 150 000-ից իջնում է (թրոմբոցիտոպենիա), թրոմբոցիտոպոետինների սինթեզն ուժեղանում է և արդյունքում թրոմբոցիտների քանակի ընկած մակարդակը կարգավորվում է։
Ակտիվացած թրոմբոցիտներից հաջողվել է ստանալ մի նոր գործոն՝ թրոմբոցիտոպոետինների ինհիբիտոր, որը ճնշող ներգործություն է ունենում թրոմբոցիտոպոեզի ինչպես միտոզի, այնպես էլ էնդոմիտոզային պրոցեսների վրա։ Արյունաստեղծման պրոցեսում նյարդային համակարգի դերը բացահայտվել է փորձարարական ճանապարհով։ Ոսկրածուծի նյարդի գրգռումը խթանում է էրիթրոպոեզը, իսկ այդ նյարդերի հատումը ընկճում։ Սիմպատիկ նյարդերի գրգռումը ուղեկցվում է լեյկոցիտոզով։ Թափառող նյարդի գրգռումն առաջ է բերում լեյկոցիտների վերաբաշխում ծայրամասային օրգանների արյան լեյկոցիտների հաշվին ներքին օրգանների արյան լեյկոցիտների քանակն ավելանում է։ Հաստատված է, որ արյունաստեղծ օրգանների նյարդերը պարունակում են ինչպես կենտրոնախույս, այնպես էլ կենտրոնաձիգ նյարդաթելեր, որոնք սկիզբ են առնում տարբեր բնույթի ռեցեպտորներից։ Նման նյարդավորման շնորհիվ ոսկրածուծն ու ավշային հանգույցները մյուս օրգան-համակարգերի հետ գտնվում են երկկողմանի կապի մեջ։ Անկասկած արյունաստեղծման վրա նյարդային համակարգի ազդեցությունը իրականանում է անուղղակի ճանապարհով` հումորալ գործոնների միջնորդությամբ, որոնց ներգործությունների վերջին օղակը համարվում են հեմոպոետինները։ Այսպիսով ամբողջ կյանքի ընթացքում արյունաստեղծման բոլոր ճյուղերի բնականոն ընթացքը ենթարկվում է խթանիչ և արգելակիչ համակարգերի հավասարակշռված նրբագույն կարգավորման, որի շնորհիվ արյան բջջային կազմն իր բազմապիսի ֆունկցիաներով պահպանվում է հարաբերական հաստատուն մակարդակի վրա։
Էլեկտրոնային նյութի սկզբնաղբյուրը ՝ Doctors.am
Նյութի էլէկտրոնային տարբերակի իրավունքը պատկանում է Doctors.am կայքին



