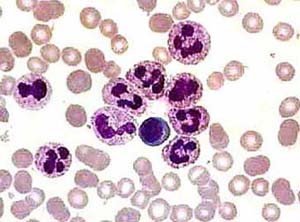
 Լեյկոցիտները կամ արյան սպիտակ բջիջներն իրագործում են բազմապիսի պաշտպանական ֆունկցիաներ։ Դրանք արյան ամենառեակտիվ բջիջներն են և արագությամբ շարժվում են վնասված կամ ախտահարված օջախը։ Լեյկոցիտների կյանքի տևողությունը, չհաշված «հիշողության» լիմֆոցիտներինը, չափազանց կարճ է` մի քանի ժամից մինչև մի քանի օր։ Անոթի վիճակում արյան 1մմ^3-ում լեյկոցիտների քանակը տատանվում է 4000-9000 (4-9-10Տ/լ) սահմաններում։ Այս քանակը կարող է ավելանալ կամ պակասել։ Լեյկոցիտների քանակի ավելացումը կոչվում է լեյկոցիտոզ որը կարող է առաջանալ ինչպես ֆիզիոլոգիական, այնպես էլ ախտաբանական պատճառներից։ Ֆիզիոլոգիական լեյկոցիտոզը զարգանում է ֆիզիկական աշխատանքի հետևանքով (մկանածին լեյկոցիտոզ), քաղցած ժամանակ կամ ուտելուց հետո (մարսողական լեյկոցիտոզ), հույզերի ազդեցությամբ (հուզական լեյկոցիտոզ), ցավից (ցավային լեյկոցիտոզ)։
Լեյկոցիտները կամ արյան սպիտակ բջիջներն իրագործում են բազմապիսի պաշտպանական ֆունկցիաներ։ Դրանք արյան ամենառեակտիվ բջիջներն են և արագությամբ շարժվում են վնասված կամ ախտահարված օջախը։ Լեյկոցիտների կյանքի տևողությունը, չհաշված «հիշողության» լիմֆոցիտներինը, չափազանց կարճ է` մի քանի ժամից մինչև մի քանի օր։ Անոթի վիճակում արյան 1մմ^3-ում լեյկոցիտների քանակը տատանվում է 4000-9000 (4-9-10Տ/լ) սահմաններում։ Այս քանակը կարող է ավելանալ կամ պակասել։ Լեյկոցիտների քանակի ավելացումը կոչվում է լեյկոցիտոզ որը կարող է առաջանալ ինչպես ֆիզիոլոգիական, այնպես էլ ախտաբանական պատճառներից։ Ֆիզիոլոգիական լեյկոցիտոզը զարգանում է ֆիզիկական աշխատանքի հետևանքով (մկանածին լեյկոցիտոզ), քաղցած ժամանակ կամ ուտելուց հետո (մարսողական լեյկոցիտոզ), հույզերի ազդեցությամբ (հուզական լեյկոցիտոզ), ցավից (ցավային լեյկոցիտոզ)։
Արյան մեջ շրջանառում է լեյկոցիտների չնչին մասը միայն։ Նրանց հիմնական զանգվածը գտնվում է հյուսվածքներում և պահեստավորված է ոսկրածուծում, մանր երակների և մազանոթների շուրջը (առպատային պահեստատեղ)։ Այդ պատճառով արյան մեջ դրանց ավելացումը կատարվում է մեծ արագությամբ և վերաբաշխողական բնույթ ունի։ Դրա շնորհիվ նույնիսկ սուր արյունահոսությունից հետո արյան լեկոցիտների քանակը մնում է անփոփոխ։ Ախտաբանական պատճառներից (բորբոքումներ, տարափոխիկ հիվանդություններ) առաջացած լեյկոցիտոզը կոչվում, է ռեակտիվ և պաշտպանական նշանակություն ունի։ Այս դեպքում լեյկոցիտների քանակն ավելանում է ախտածին գործոնների ազդեցությամբ լեյկոպոեզի խթանման շնորհիվ, և արյան մեջ ի հայտ են գալիս նաև լեյկոցիտների երիտասարդ ձևեր։ Ռեակտիվ լեյկոցիտոզը ժամանակավոր բնույթ ունի և տեղի է ունենում տասնյակ հազարների սահմաններում։ Լեյկոցիտների քանակն ավելանում է նաև սպիտակարյունության ժամանակ (լեյկոզ կամ լեյկեմիա)։ Վերջինիս դեպքում լեյկոցիտների քանակի ավելացումը հասնում է հարյուր հազարների և կայուն բնույթ է կրում։ Դա ոսկրածուծի ծանր հիվանդություն է և ենթակա է բուժման։
Լեյկոցիտների քանակի պակասը լեյկոպենիան, ռադիոակտիվ ճառագայթման մակարդակի բարձրացման հետ կապված ներկայումս ավելի հաճախ է հանդիպում։ Այդ է պատճառը, որ նախկինում արյան 1մմ3 ծավալում լեյկոցիտների բնականոն քանակի ստորին սահմանը (6000) այժմ համեմատաբար ավելի փոքը թիվ է (4000): Ճառագայթային հիվանդության ժամանակ լեյկոպոեզի ախտահարումն ուղեկցվում է ծանր լեյկոպենիայով, որի համար երբեմն դիմում են ոսկրածուծի փոխպատվաստման։ Ցիտոստատիկ դեղանյութերի ազդեցությամբ լեյկոպոեզի ընկճումը նույնպես հանգեցնում է լեյկոպենիայի։ Լեյկոպենիայի հետևանքով օրգանիզմի դիմադրողականությունը վարակների հանդեպ ընկնում է։ Երբ լեյկոցիտների քանակը արյան 1մմ3-ում հասնում է 500-ի (!) մահը դառնում է անխուսափելի։
Լեյկոցիտներն անգույն բջիջներ են և ուսումնասիրելու համար նրանց ներկում են։ Ներկելուց պարզվում է, որ նրպնց մի մասի ցիտոպլազման պարունակում է հատիկներ, իսկ մյուս մասինը հատիկներ չի պարունակում։ Հատիկներ պարունակող լեյկոցիտները կոչվում են հատիկավորներ (գրանուլոցիտներ), իսկ չպարունակողները՝ ոչ հատիկավորներ (ագրանուլոցիտներ)։ Գոյությունն ունեն 3 տեսակ հատիկավոր լեյկոցիտներ` բազոֆիլներ, էոզինոֆիլներ և նեյտրոֆիլներ։ Առողջ մարդու արյան մեջ կան ոչ միայն հասուն նեյտրոֆիլներ՝ սեգմենտակորիզներ, այլ նաև ցուպիկակորիզավորներ, իսկ երբեմն նաև՝ մետամիելոցիտներ
կամ երիտասարդ ձևեր։ Ոչ հատիկավոր լեյկոցիտները երկու տեսակ են՝ մոնոցիտներ և լիմֆոցիտներ։ Բոլոր այս բջիջներն ունեն իրենց բնորոշ պաշտպանական ֆունկցիաները և նրանցից յուրաքանչյուրի քանակի ավելացումը տեղի է ունենում համապատասխան գործոնի ազդեցությամբ։ ՈՒստի կլինիկական կարևոր արժեք ունի ոչ միայն արյան լեյկոցիտների բացարձակ քանակի, այլ նաև դրանց տարբեր տեսակների տոկոսային հարաբերության որոշումը, որը կոչվում է լեյկոցիտային բահաձև (լեյկոֆորմուլ կամ լեյկոգրամ)։ Լեյկոգրամի որոշումն ունի ախտորոշիչ արժեք։
Օրգանիզմի պաշտպանական ռեակցիաները, որոնց մեջ ընդգրկվում են նաև լեյկոցիտները, իրագործվում են երկու մեխանիզմներով՝ ոչ յուրահատուկ, ոչ սպեցիֆիկ և յուրահատուկ՝ սպեցեֆիկ մեխանիզմներով։ Ոչ սպեցիֆիկ են կոչվում այն մեխանիզմները, որոնք գործում են բնորոշ ներգործություն չունեցող ազդակների հանդեպ և արդյունավետ են ցանկացած օտարածին նյութի նկատմամբ։ Այս մեխանիզմներին են պատկանում մի կողմից արտաքին ու ներքին միջավայրի պատնեշները, իսկ մյուս կողմից՝ ներքին միջավայրի հումորալ ու բջջային մեխանիզմները։ Արտաքին միջավայրի պատնեշներն են համարվում մաշկի վերնամաշկը և լորձաթաղանթի էպիթելը, իսկ ներքին միջավայրինը՝ հյուսվածքաարյունային պատնեշները։
Ներքին միջավայրի հումորալ մեխանիզմներն իրականանում են արյան սպիտակուցային նյութերի միջոցով, որոնցից կարևորվում են երկու համակարգեր`պրոպերդինային և կոմպլեմենտային։ Կոմպլեմենտային համակարգը իր մեջ ընդգրկում է պլազմայի 11 սպիտակուցներ (C1 C2 C3 և այլն)։ Ոչ սպեցիֆիկ մեխանիզմներին մասնակցում են նաև հյուսվածքային հեղուկում և արյան պլազմայում գտնվող լեյկինները, պլակինները, բետալիզինը։ Ոչ սպեցիֆիկ պաշտպանության բջջային մեխանիզմն իրականանում է բորբոքային ռեակցիաների և ֆագոցիտոզի ձևով։ Վերջինս կատարվում է միկրոֆագերի (նեյտրոֆիլներ) և մակրռֆագերի կողմից։ Ներկայումս մակրոֆագերը, մոնոցիտները, հիստիոցիտները, լյարդի կուպֆերյան բջիջները միավորվում են օրգանիզմի պաշտպանական համակարգերից մեկի` մոնոնուկլեար ֆագոցիտային համակարգի (ՄՖՀ) մեջ։ Ֆագոցիտոզի կարող են ենթարկվել ցանկացած օտարածին մարմիններ, որի համար այն դասվում է ոչ յուրահատուկ մեխանիզմների շարքը։ Սակայն ոչ յուրահատուկ պաշտպանական հարմարանքները հաճախ ընթանում են յուրահատուկ մեխանիզմներին զուգընթաց, լրացնում նրանց և նպաստում պաշտպանական ռեակցիաների զարգացման բնականոն ընթացքին։
Սպեցիֆիկ պաշտպանական մեխանիզմները գործում են խիստ ընտրողաբար, որոշակի օտարածին ազդակի հանդեպ, նրա նկատմամբ մշակելով համապատասխան հակազդող ներքին գործոն։ Դրա հիմքում ընկած է իմունիտետը։ Իմունիտետն օրգանիզմի պաշտպանական ռեակցիան է օտար գենետիկական ինֆորմացիա ունեցող ագդակների վիրուսների, միկրոօրգանիզմների, մակաբույծների, բջիջների, հյուսվածքների և օրգանների հանդեպ։ Իմունիտետի գործող բջիջները լիմֆոցիտներն են, որոնք համագործակցում են մյուս լեյկոցիտների հետ ։ Այդ միասնական գործունեության հետ միասին յուրաքանչյուր լեյկոցիտ ունի նաև իրեն բնորոշ որոշակի ֆունկցիաները։
Բազոֆիլներ։ Ամենափոքրաթիվ լեյկոցիտներն են` 0-1%: Բազոֆիլները և պարարտ բջիջներն ունեն ընդհանուր ծագում։ Պարարտ բջիջներին անվանում են նաև հյուսվածքային բազոֆիլներ։ Այս երկու բջիջներին միավորում են հեպարինոցիտների խմբում։ Դրանց ցիտոպլազման պարունակում է հեպարին, հիստամին և կենսաբանական այլ ակտիվ նյութեր, որոնց բազոֆիլները կլանում են հյուսվածքներից կամ իրենք են սինթեզում, իսկ անհրաժեշտության դեպքում ենթարկում սեկրեցիայի։ Հեպարինը հակամակարդիչ նյութ է, իսկ հիստամինը անոթալայնիչ։
Հեպարինոցիտները պատում են լյարդի (որտեղ երակային արյունը դանդաղ է հոսում) և թոքերի (որտեղ թրոմբոցիտների քանակը մեծ է) մանր անոթները և հեպարինի միջոցով կանխում արյան ներանոթային մակարդման վտանգը։ Հիստամինը բորբոքումների ժամանակ նպաստում է հյուսվածքների վերականգնման, նոր մազանոթների գոյացման, հյուսվածքների սնուցման և վերքերի ապաքինման պրոցեսներին։
Ալերգիկ ռեակցիաների ժամանակ բազոֆիլները արձակում են «էոզինոֆիլների քեմոտաքսիսի գործոն», որի ձգումով էոզինոֆիլները շարժվում են դեպի բազոֆիլների կուտակման վայրը և քայքայում նրանց պարունակած հիստամինի մեծ քանակությունը, որն օրգանիզմի համար անոթալայնիչ մեծ վտանգ է ներկայացնում։ Բազոֆիլները, ինչպես և մյուս լեյկոցիտները, օժտված են քեմոտաքսիսի և գաղթի (միգրացիա) ունակությամբ;Նրանց քեմոտաքսիսին նպաստում են լիմֆոցիտներից արտադրվող լիմֆոկինները: Օրգանիզմի գերզգայուն վիճակներում բազոֆիլներից արձակվում է նաև «անաֆիլաքսիայի դանդաղ հակազդող նյութ» որն առաջացնում է հարթ մկանների կծկում։ Հետաքրքիր է նշել, որ բազոֆիլները սինթեզում են նաև «թրոմբոցիտներն ակտիվացնող գործոն», որը որոշակի պայմաններում նպաստում է թրոմբոցիտների ագրեգացիային և նրանց պարունակության արտամղմանը։Բազոֆիլները իրենց ֆունկցիան դրսևորում են հատիկազրկման (դեգրանուլացիա) ճանապարհով։ Հատիկազրկման հզոր գործոն է համարվում Е իմունոգլոբուլինը։
Քեմոտաքսիսի շնորհիվ էոզինոֆիլները մի շարք մակաբույծ որդերի նկատմամբ իրագործում են նաև իրենց հակահելմինթային ֆունկցիան։ Նրանք ֆիքսվում են, օրինակ, շիստոսոմուլի վրա, սեփական հատիկների պարունակությամբ քայքայում մակաբույծի ծածկող հյուսվածքները, թափանցում հյուսվածքների մեջ և ոչնչացնում նրան (շիստոզոմատոզը տարածված է արևադարձային երկրներում)։
Ալերգիկ ռեակցիաների ժամանակ էոզինոֆիլները կուտակվում են ալերգիայի ենթակա օրգաններում (բրոնխիալ ասթմայի ժամանակ՝ օրինակ, շուրջբրոնխային հյուսվածքներում) և քայքայում ալերգիային նպաստող գործոնները։ Մարդկանց էոզինոֆիլները ավելի շատ կուտակվում են արտաքին միջավայրի հետ շփվող օրգաններում (մաշկ, թոքեր, մարսողական և միզասեռական օրգաններ)։ Արյան էոզինոֆիլների քանակի ավելացումը կոչվում է էոզինոֆիլիա: Այն վկայում է մակաբույծ որդերի կամ ալերգիական ռեակցիաների առկայության մասին։
Նեյտրոֆիլներ։ Կազմում են արյան լեյկոցիտների ամենամեծ խումբը 50-70%: Ըստ հասունացման աստիճանի առողջ արյան մեջ հանդիպում են մետամիելոցիտներ, կամ երիտասարդ ձևեր (0-1%), ցուպիկակտիզավորներ (1-5%) և սեգմետակտի զավորներ (45-65%), իսկ հիվանդագին դեպքերում` նաև միելոցիտներ։ Արյան մեջ շրջանառում են նեյտրոֆիլների ընդամենը 1%-ը իսկ մնացածը գտնվում են օրգաններում և հյուսվածքներում։ Նեյտրոֆիլների ցիտոպլազմայում գոյություն ունեն տիպի նեյտրոֆիլ հատիկներ; Առաջին խումբ հատիկները ֆունկցիոնալ իմաստով լիզոսոմներ են, որոնք պարունակում են բազմապիսի ֆերմենտներ լիզոցիմ, որը քայքայում է բակտերիաների թաղանթները, կատիոնային սպիտակուցներ, որոնք խանգարում են միկրոօրզանզմների շնչառության և աճի բնականոն ընթացքը; պրոտեազներ, հիդրոլազներ և այլն։ Այս ֆերմենտների ներգործությամբ է իրականանում ֆագոցիտոզի ենթարկված մարմինների ներբջջային մարսողությունը։ Երկրորդ տիպի հատիկները պարունակում են դարձյալ լիզոցիմ, վիտամին В 12-ի փոխադրիչներ, իսկ երրորդ տիպինը թթու գլիկոզամինագլիկաններ, որոնք հյուսվածքներում մասնակցում են բազմացման, աճի և վերականգնման պրոցեսների կարգավորմանը։ Երկրորդ և երրորդ տիպի հատիկները սեկրետոր օրգանոիդ են և գործում են ֆագոցիտոզից անկախ: Դրա շնորհիվ նեյտրոֆիլները դասվում են կենսաբանական ակտիվ նյութեր արտադրող բջիջների շարքը։
Այնուամենայնիվ նեյտրոֆիլների հիմնական պաշտպանական ֆունկցիան ֆագոցիտոզն է։ Բորբոքված, վնասված, կամ վարակի ենթարկված հյուսվածքներում արտադրվում են քեմոտաքսիկ և անոթալայնիչ նյութեր, որոնք որոշում են նեյտրոֆիլների գաղթի ուղղությունը։ Այդ նյութերը մեծացնում են նաև մազանոթների անդրբջջային թափանթելիությունը և, առանց մազանոթի պատը վնասելու, նեյտրոֆիլներն իրենց կեղծ ոտիկներով շարժվում են դեպի ախտահարված հյուսվածքները։ Դրանք համարվում են լեյկոցիտների ավանգարդը, որովհետև առաջինն են հասնում բորբոքային օջախներ։ Նրանց շարժման արագությունը հասնում է 40մկմ/րոպե։ Մեկ նեյտրոֆիլը կարող է կլանել 20-30 բակտերիա, որի համար էլ կոչվում են միկրոֆագեր։ Նեյտրոֆիլների մանրէասպան հատկությունը պայմանավորված է ոչ միայն լիզոսոմների ֆերմենտների ներգործությամբ, այլև. նրանց այսպես կոչված «նյութափոխանակության բռնկումով»։ Այս երևույթն սկսվում է մանրէի և նեյտրոֆիլի թաղանթների հպումից 30-60 վրկ հետո: Մեծանում է նեյտրոֆիլի Օշ-ի յուրացման աստիճանը, առաջանում են Օշ իոններ, Н202 և այլ ակտիվ նյութեր, որոնք մասնակցում են մանրէնրի թաղանթների քայքայմանը։ Նեյտրոֆիլների կողմից արտադրված ինտերֆերոնը ոչնչացնում է վիրուսներին։ Որոշ դեպքերում բակտերիաների թույներից ոչնչանում են իրենք` նեյտրոֆիլները։
Նեյտրոֆիլների քանակի ավելացումը կոչվում է նեյտրոֆիլոզ։ Հիվանդագին դեպքերում (թարախային բորբոքումներ, տարափոխիկ հիվանդություններ և այլն) այն ունի ռեակտիվ բնույթ և զարգանում է նեյտրոֆիլների երիտասարդ ձևերի հաշվին։ Նեյտրոֆիլների այդ ռեակցիան կոչվում է ձախ թեքումով նեյտրոֆիլոզ։ «Ձախ» է կոչվում, որովհետև լեյկոցիտային բանաձևի աղյուսակում երիտասարդ նեյտրոֆիլները նշված են նրա ձախ կողմում, իսկ հասուն և ծերացած սեգմենտակորիզների հաշվին զարգացող նեյտրոֆիլոզը կոչվում է «աջ» թեքումով։ Նեյտրոֆիլոզի ձախ թեքման չափը որոշում են ռեգեներացիայի ցուցանիշի օգնությամբ։ Դա երիտասարդ ձևերի (միելոցիտ,մետամիելոցիտ և ցուպիկակորիզ) գումարի և սեգմենտակորիզավորների քանակի հարաբերությունն է։ Առողջ օրգանիզմում այն հավասար է 0,05-0,1 ի, իսկ ծանր ինֆեկցիաների և բորբոքումների ժամանակ հասնում է 1-2-ի։ Երիտասարդ նեյտրոֆիլների առկայությունը մի կողմից վկայում է հիվանդության ընթացքի ծանրության, իսկ մյուս կողմից օրգանիզմի պաշտպանական ռեակցիաների հուսալիության մասին։ Հակառակ դեպքում, երբ նեյտրոֆիլոզը բացակայում է, կամ նրանց ֆունկցիոնալ ունակություններն անբավարար են, հիվանդությունը կարող է անցանկալի ելք ունենալ։
Ինֆեկցիոն հիվանդությունների ընթացքին նեյտրոֆիլների նման մասնակցությունը վկայում է որ մակրոֆագերի և լիմֆոցիտների հետ միասին օրգանիզմի սպեցիֆիկ պաշտպանական մեխանիզմներում նեյտրոֆիլներն ունեն իրենց որոշակի դերը։
Մոնոցիտներ։ Կազմում են արյան լեյկոցիտների 2-9%-ը։ Համարվում են մոնոնուկլեար ֆագոցիտային համակարգի (ՄՖՀ) կենտրոնական օղակը, որովհետև օժտված են ֆագոցիտային և մանրեասպան բարձր ունակությամբ։ Արյունից մոնոցիտները գաղթում են մարմնի հյուսվածքներն ու խոռոչները, որտեղ կատարվում է նրանց տարբերակումը։ Դրանք չափերով մեծանում են, հարստանում լիզոսոմներով, որոնց մեջ ավելանում է նաև ֆերմենտների քանակը։ Մասնագիտացած այս մոնոցիտներին հաճախ անվանում են մոնոցիտ-մակրոֆագեր։
Բորբոքային օջախներում ինֆեկցիայի չենթարկված հյուսվածքներում մոնոցիտ-մակրոֆագերը կարող են կլանել մինչև 100 մանրէ։ Նրանց ֆագոցիտային ունակությունն առավելագույն չափի է հասնում թարախային բորբոքային օջախներում, որտեղ pH-ը թթվային է։ Նման pH-ի պայմաններուսմ նեյտրոֆիլները կորցնում են իրենց ֆագոցիտային հատկությունը, իսկ մակրոֆագները կլանում են և. միկրոբները և մահացած նեյտրոֆիլները։
Ֆագոցիտոզն ապահովող էներգիան մակրոֆագերն ստանում են էներգիայի փ-խանակության թթվածնային փուլում անջատված էներգիայից։ Ֆագոցիտոզի ընթացքում ուժեղանում է Օ2-ի յուրացումը, օքսիդացման հետևանքով անջատվում են Օշ-,HշՕշ և այլ ակտիվ նյութեր, որոնք մանրէասպան հատկություն ունեն։ Թթվածնից զուրկ պայմաններում, օրինակ, թարախակույտում, ֆագոցիտոզը շարունակում է անթթվածնային ճեղքման էնեիգիայի հաշվին։ Մակրոֆագերը կլանում են նաև ծերացած և վնասված բջիջները, մաքրում հյուսվածքները, նպաստելով առողջ և վնասված հյուսվածքների վերականգնման պրոցեսներին։ Այս գործունեության համար նրանց անվանում են «օրգանիզմի հավաքարարներ»։ Մարդկանց մակրոֆագերն արտադրում են ավելի քան 100 կենսաբանական ակտիվ նյութեր` ինտերլեյկին-1, կախեքսին,
լիզոցիմ, ինտերֆերոն, կոմպլեմենտ, էլաստազա, պլազմոգեն, ֆիբրոգեն և այլն։ Ինտերֆերոնով է պայմանավորված մակրոֆագերի հակաուռուցքային, իմունոկարգավորման, հակավիրուսային և. այլ ազդեցություններ։ Ֆիբրոգենն արագացնում է կոլագենի սինթեզը և նպաստում ֆիբրոզ հյուսվածքի ձևավորմանը: Ինտերլեյկին I-ը խթանում է Т-լիմֆոցիտների և օստեոբլաստների բազմացումը, իսկ կախեքսինը ուռուցքային բջիջները ենթարկում է. մեռուկի (նեկրոզ)։ Վերջին երկու նյութերը միաժամանակ գրգռում են նաև ջերմագոյացման կենտրոնը և բարձրացնում օրգանիզմի ջերմությունը։
Ֆագոցիտոզի ընթացքում մակրոֆագերը անտիգեններից մշակում են հատուկ նյութեր` իմունոգեններ, որոնց անմիջական մասնակցությամբ սկիզբ են առնում իմունոլոգիական բարդ ռեակցիաներ։ Իմունիտետի զարգացման մեխանիզմում մոնոցիտները սերտ համագործակցության (կոոպերացիա) մեջ են մտնում լիմֆոցիտների հետ։
 Լիմֆոցիտներ։ Կազմում են լեյկոգրամի 20-40%: Օրգանիզմում լիմֆոցիտների ընդհանուր զանգվածն անհամեմատ մեծ է 41,5կգ։ Լիմֆոցիտներն անընդհատ շրջում են մի հյուսվածքից մյուսը։ Ի տարբերություն մյուս լեյկոցիտների, նրանք հյուսվածքից կրկին վերադառնում են արյուն և ապա գաղթում դեպի, մի այլ հյուսվածք։ Լիմֆոցիտների մի մասը իմունոլոգիական հիշողություն ունեցողները) երկարակյաց են և կարող են ապրել 10-20 և ավելի տարի։
Լիմֆոցիտներ։ Կազմում են լեյկոգրամի 20-40%: Օրգանիզմում լիմֆոցիտների ընդհանուր զանգվածն անհամեմատ մեծ է 41,5կգ։ Լիմֆոցիտներն անընդհատ շրջում են մի հյուսվածքից մյուսը։ Ի տարբերություն մյուս լեյկոցիտների, նրանք հյուսվածքից կրկին վերադառնում են արյուն և ապա գաղթում դեպի, մի այլ հյուսվածք։ Լիմֆոցիտների մի մասը իմունոլոգիական հիշողություն ունեցողները) երկարակյաց են և կարող են ապրել 10-20 և ավելի տարի։ Լիմֆոցիտներն իմունիտետի գործող բջիջներն են։ Նրանք վնասազերծում են օրգանիզմ թափանցած օտար գենետիկական տեղեկատվություն ունեցող մանրէներ, մակաբույծներ, բջիջներ, հյուսվածքներ և օրգաններ։ Տարբերում են 3 տիպի լիմֆոցիտներ` T-լիմֆոցիտներ, B-լիմֆոցիտներ և զրոյական լիմֆոցիտներ։ Վերջիններս անհրաժեշտության դեպքում տարբերակվում են Т- կամ B-լիմֆոցիտների: T-լիմֆոցիտների տարբերակումը տեղի է ունենում ուրցագեղձում (thumus, որի սկզբնատառից ծագում Է այս բջիջների անվանումը)։ Թռչունների B-լիմֆոցիտները տարբերակվում են հատուկ ավշային օրգանում` bursa Fabricius (bursa բառի սկզբնատառից ծագում Է B-լիմֆոցիտների անվանումը)։ Կաթնասուների և մարդկանց օրգանիզմում այդ օրգանը բացակայում Է։ Նրանց B-լիմֆոցիտները մասնագիտանում են մարսողական օրգանների` որդանման ելուստի,պեյերյան բծերի, քիմքի և ըմպանի ավշային հանգույցներում։ T-լիմֆոցիտների քանակը կազմում Է արյան լիմֆոցիտների շուրջ 70%-ը։
T-լիմֆոցիտներն առաջանում են ոսկրածուծում։ Դեռևս սաղմնային զարգացման շրջանում նրանք արյան միջոցով փոխադրվում են ուրցագեղձ, որտեղ տարբեր բնույթի անտիգենների նկատմամբ ձեռք են բերում համապատասխան ընկալիչներ։ Վերջիններս դասավորվում են թաղանթի վրա։ Ուրցագեղձից T-լիմֆոցիտները փոխադրվում են ծայրամասային ավշային հանգույցները։ Այս լիմֆոցիտները «նստակյաց» չեն և արյան ու ավշի միջոցով անընդհատ շրջում են օրգանիզմի տարբեր մասերում։ Հյուսվածքների մեջ թափանցած օտար մարմինները մակրոֆագերի կողմից ենթարկվում են ֆազոցիտոզի, որի ընթացքում մշակվում են իմունոգեններ։ Մակրոֆագերը դրանցից անջատված անտիգենները հասցնում են T-լիմֆոցիտներին, որոնց ընկալիչները միանում են համապատասխան անտիգենների հետ։ Այս միացությունը խթանում է T-լիմֆոցիտներին, որոնք սկսում են բազմանալ,առաջացնելով T-լիմֆոցիտների տարատեսակներ։ Տարբերում են՝ T-ճնշողներ (սուպրեսորները), Т-օգնականներ (հելպերներ), T-սպանիչներ (քիլերներ), T-ակտիվացնողներ (ամպլիֆայերներ)։ T-սուպրեսորները չափավորում են B-լիմֆոցիտների ուժեղացած ֆունկցիան, իսկ T-հելպերները փոխներգործում են B-լիմֆոցիտների հետ և նրանց դարձնում պլազմատիկ բջիջներ։ T-ամպլիֆայերը ակտիվացնում են T-քիլեր լիմֆոցիտներին, որոնք սինթեզված հատուկ նյութերի` լիմֆոկինների միջոցով քայքայում են, օրինակ, փոխպատվաստած օտար գենետիկական տեղեկատվություն ունեցող բջիջներ, հյուսվածքներ և օրգաններ։ Դրա շնորհիվ վերջիններս մերժվում են օրգանիզմի կողմից։ T-լիմֆոցիտները քայքայում են նաև քաղցկեղի բջիջները, մուտացիայի ենթարկված բջիջները։ Յուրաքանչյուր Т-լիմֆոցիտ քայքայում է մեկ բջիջ։ Իմունիտետի այս ձեր իրականանում է արյան բջիջների` T-լիմֆոցիտների անմիջական մասնակցությամբ, ուստի այն կոչվում է բջջային իմունիտետ։ Պաշտպանական այս յուրահատուկ ռեակցիաների ընթացքում ձևավորվում են նաև հիշողության Т-լիմֆոցիտները, որոնք առանց կիսվելու պահպանվում են տարբեր հյուսվածքներում և նույն անտիգենին հանդիպելիս ապահովում համապատասխան իմունոլոգիական ռեակցիան։
B-լիմֆոցիտները նույնպես առաջանում են ոսկրածուծում, տարբեր անտիգենների նկատմամբ ձեռք բերում .համապատասխան ընկալիչներ, և փոխադրվում ավշային հանգույցների, փայծաղի, պեյերյան բծերի մեջ։ Այստեղ թափանցած մանրէների անտիգենների առկայության մասին տեղեկատվություն նրանք ստանում են T-հելպերներից, որի շնորհիվ ակտիվանում են, փոխադրվում մոտակա ավշային հանգույց և բազմանում։ Բազմացած այդ բջիջների մի մասը դառնում է հիշողության B-լիմֆոցիտներ, որոնք առանց կիսվելու երկար տարիներ պահպանվում են հյուսվածքներում սպասելով կրկին հանդիպելու համապատասխան անտիգենի։ Իսկ մյուս մասը տարբերակվում է պլազմատիկ բջիջների, որոնք սինթեզում են անտիգենին համապատասխան հակամարմիններ՝ իմունոգլոբուլիններ։ Վերջիններս արյան և ավշի միջոցով (հումորալ ճանապարհով) հասնում են վարակի ենթարկված օջախը և ոչնչացնում հարուցիչները։ Իմունիտետի այս ձևը անվանում են հումորալ։
Լիմֆոցիտները մասնակցում են նաև կրեատոր մակրոմոլեկուլների փոխադրմանը։ Դրանով նրանք մասնակցում են օրգանների փոխադարձ կապի և օրգանիզմի ամբողջականության պահպանմանը։
Էլեկտրոնային նյութի սկզբնաղբյուրը ՝ Doctors.am
Նյութի էլէկտրոնային տարբերակի իրավունքը պատկանում է Doctors.am կայքին